Some Research highlights
Exceptionally preserved fossil deposits and their evolutionary significance are the major focus of my research. These important windows on the life of the past provide much more data than normal shelly fossil deposits. My research on these fossils has involved collaborations with students and postdocs as well as colleagues across the world (see publications list) whose fundamental contributions are gratefully acknowledged.
I took part in explorations which demonstrated that the original Cambrian Burgess Shale locality in British Columbia is far from unique, and that a large number of adjacent localities also yield exceptionally preserved Cambrian fossils. Burgess Shale-type fossils have since been found almost worldwide and remain a major focus of palaeontological research. It is now known that many types of exceptional preservation are not confined to unique or even unusual settings.
I have applied a combination of new experimental approaches, and chemical and mineralogical analyses, to investigate the fossilization of soft-tissues. This has involved research on the replication of soft-tissues in minerals such as apatite and pyrite (the Medusa effect). Geochemical and isotopic studies of fossils and sediments from Beecher’s Trilobite Bed (Ordovician, New York State) and the Hunsrück Slate (Devonian, Germany) revealed the conditions which gave rise to the pyritization of soft-bodied fossils. Experiments on the underlying biogeochemical mechanisms provided insights into the rates, conditions and controls involved in the fossilization of soft-tissue morphology. Replication is most rapid in calcium phosphate minerals, which preserve the highest fidelity of detail.

The trilobite Triarthrus, preserved in pyrite, from the Ordovician of Upstate New York.
Some organic compounds, such as melanin, have survived almost unchanged in fossils at least as old as Mesozoic but most organic materials are subject to diagenesis. Chitin is present in insects as old as 25 million years and traces occur in much older rocks, but it is not the original chemistry that survives in most fossil arthropods—the cuticle has been altered to an aliphatic composition through diagensis. The transformation of insect and plant tissues to a kerogen-like substance can be achieved in the laboratory. The fossilization of organic materials is a result of the diagenetic transformation of lipids in the organism itself by in situ polymerization. More recently research has revealed the conditions favorable to the preservation of proteinaceous tissues in fossil hard parts such as dinosaur bone and shown that they retain information indicative of relationships and phylogenetic position, modes of biomineralization, and metabolic rate.
The exceptionally preserved Ordovician fauna from the Fezouata formations of Morocco showed that machaeridians, a mystery since 1857, previously only known from shelly plates, were armored annelids. The Fezouata formations have yielded a diverse Burgess Shale-type fauna including a giant filter feeding radiodont Aegirocassis and opened up a new window on the Great Ordovician Biodiversification Event.

The arthropod Furca from the Ordovician Fezouata shales, Morocco.
High fidelity 3D reconstructions of rare, soft-bodied Silurian fossils from Herefordshire in England can be rotated and ‘dissected’ on the computer screen. The Herefordshire Lagerstätte has yielded radiolarians, sponges, a polychaete worm, aplacophoran-like molluscs, a platyceratid gastropod, orthoconic nautiloids, several arthropods including trilobites, a marrellomorph, a pycnogonid, a stem-group chelicerate, ostracodes, a barnacle and a phyllocarid, a brachiopod, several echinoderms, and graptolites. Many of these Herefordshire animals provide critical insights into the early evolutionary history of the groups to which they belong.
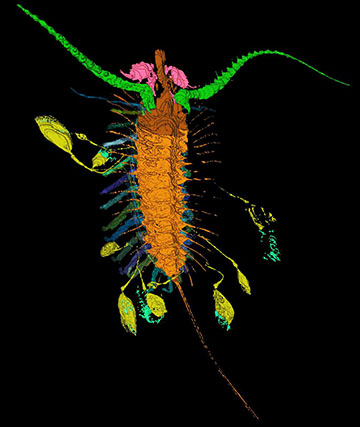
The remarkable arthropod Aquilonifer from the Herefordshire biota with juveniles attached.
Novel methods of interpreting color in fossils have resulted in unforeseen insights into the ecology of extinct animals. Feathers are preserved as a result of the decay resistance of melanosomes. The microstructure of fossil feathers to be compared to that in living birds. The analysis of samples of a specimen of Anchiornis from the Jurassic of China led to the first scientific reconstruction of color patterns in the plumage of an entire dinosaur. Reconstructions of color in fossil insects, particularly lepidopterans and beetles, have also proved possible.

A chrysomelid beetle from the Eocene of Eckfeld, Germany. The color is structural in origin.
The famous Bertie Group deposits of upper New York State and southern Ontario are the source of the Ciurca collection of eurypterids at the Yale Peabody Museum. Other rare forms present alongside the eurypterids including a large disc-like hydrozoan with a float up to 17 cm in diameter and a strange pterobranch, related to graptolites, which Samuel Ciurca called Ezekiel’s Wheel.

A giant complete specimen of the eurypterid Acutiramus from the Ciurca collection.